 Theory of Stuttering – Blog
Theory of Stuttering – Blog
Theory of Stuttering – Blog
The basal ganglia have been suspected to play a key role in the causation of stuttering for a long time, especially since Per Alm (2004) proposed his model in which he assumes an “impaired ability of the basal ganglia to produce timing cues for the initiation of the next motor segment in speech” (p. 325). A role of the basal ganglia in stuttering is suggested also by functional brain imaging studies in which overactivations were found in the striatum (Braun et al., 1997; Fox et al., 1996; Ingham et al., 2003) and in the substantia nigra (Metzger et al., 2018; Watkins et al., 2008; Wu et al., 1997) .
Some neuroimaging studies have reported altered structural properties of the basal ganglia in stutterers, but these results are in part inconsistent (see Table 1 in Montag et al., 2019). However, in a recent MRI investigation of 33 stutterers and 34 controls, Neef et al. (2018) found the right nucleus accumbens to be larger in participants who stuttered compared to controls. This result has now been confirmed by Montag et al. (2019) in a study involving 36 stutterers and 34 controls: the authors found increased gray matter volume in the right ventral striatum in the stutterers compared to the controls.
It is of theoretical importance how the authors of both studies interpret their findings. Neef and colleagues refer to recent theories of basal ganglia function suggesting that the nucleus accumbens is a “motivation-to-movement interface” (p. 84). Corroborating this view, Montag and colleagues write: “The location of our finding is noteworthy, as previous work on basal ganglia involvement in stuttering has mainly focused on motor function. The ventral striatum region belongs to the limbic subdivision of the basal ganglia circuit which is involved in reward and motivational functions” (p. 4).
One purpose of this blog is to discuss whether my theory of stuttering is consistent with new empirical findings, and I think the theory is consistent with the insight that the role of the basal ganglia in stuttering is not related to motor control as such, but rather to the cause of (or the reason for) motor action, namely to motivation and reward. I have never believed that a deficit in the basal ganglia function causes stuttering, but that invalid error signals in the monitoring system cause interruptions of speech flow. These signals that suddenly inhibit motor execution seem to originate from the left cerebellum (see Section 2.2. in the main text ).
Nevertheless, Alm’s (2006, 2010) theory which explains stuttering in the framework of Goldberg’s (1985) dual premotor model is still a fruitful approach. The problem with Goldberg’s model is that it differentiates between (1) automatic, attention-independent motor actions controlled by the medial premotor system (SMA, basal ganglia) and (2) motor actions depending on attention and sensory feedback which are controlled by the lateral premotor system (lateral premotor cortex, cerebellum; compare Alm, 2010).
It is, however, known that the SMA controls voluntary, internally initiated action (it may be more or less automatized), and that the cerebellum plays an important role in motor sequence learning, i.e., in the automatization of sequential motor behavior. It may therefore be more appropriate to assume that voluntary, internally initiated action is controlled by the medial premotor system, whereas involuntary, externally initiated, or feedback-dependent action is controlled by the lateral premotor system. Both kinds of motor action is associated with attention: voluntary action with selective (top-down) attention, involuntary motor action with non-selective (bottom-up) attention.
Like other complex and automatized human behavior, peaking obviously includes both kinds of motor action and both control systems: Speaking is indeed in part controlled by the will: we decide whether to speak or not, we choose content words and emphasize important words. But most other aspects of spontaneous speech in one’s native language run automatic and involuntary: inserting function words, observing syntax and, and grammar rules, linguistic stress, and even more the production of phonemes and phoneme transitions. This latter, most automatic aspect of speech is impaired in stuttering.
The automatic aspects of speech, e.g., rhythmic syllable starts might indeed be feedforward controlled by the medial premotor system. However, the flow of speech is, in a negative sense, depending on the self-monitoring system, since speech errors usually require an interruption for repair. The self-monitoring of overt (and audible) speech depends on auditory feedback (Lind et al., 2014; see also this blog entry). Error detection and -response seems to be a function of the lateral premotor system, particularly of the cerebellum (e.g., Zheng et al., 2013; see also Section 2.2 in the main text). Poor processing of the sensory feedback of speech, after my theory, results in invalid error signals causing interruptions of speech flow, against the speaker’s will and (virtually) for no clear reason: The execution of a speech motor program is suddenly inhibited. The medial premotor system then behaves rather normally when it tries to overcome the inhibitions – but this causes the observable symptoms of stuttering: repetitions, prolongations, the tension in a block (see Section 2.1 in main text and PDF).
This view is consistent with the observation that dopamine-blocking drugs like haloperidol, by damping the activity of the basal ganglia and, by that, of the medial premotor system, reduce the severity, but hardly the frequency of stuttering symptoms. So Alm (2004, p. 337) writes: “The drug seems to exert its main effect on the severity of stuttering behavior and not so much on the frequency of stuttering”, Similarly, Stager et al. (2005) found that pimozide (a dopamine antagonist) shortened the average duration of dysfluencies, but did not reduce the number of disfluencies per minute.
So I propose to simply reverse Alm’s theory: Not the function of the medial premotor system is impaired in stuttering, but the function of the lateral premotor system is impaired. This idea is not new: Watson and Freeman (1997) discussed studies of regional cerebral blood flow (rCBF) and studies of speech motor performance in which acoustic laryngeal reaction time (LRT) was measured as a function of the complexity of the required response, e.g., a word or sentence. They wrote: “The rationale for the LRT studies arose, in part, from Goldberg’s (1985) discussion of medial and lateral premotor systems […] Relations between resting rCBF anomalies and speech motor performance deficits are consistent with predictions of Goldberg’s (1985) model regarding defects in the lateral premotor system ” (pp. 343/44).
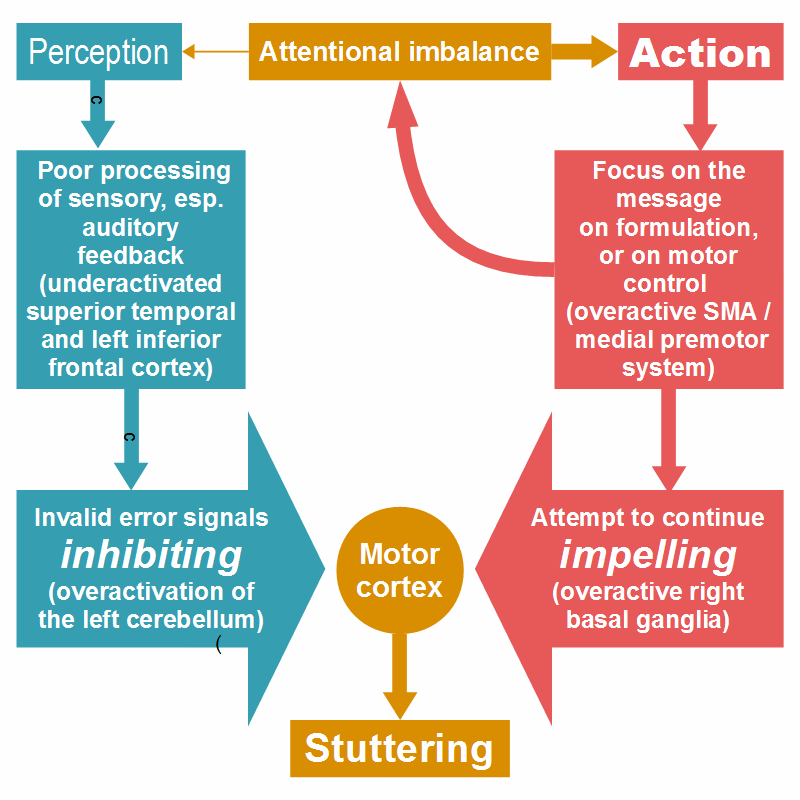
Figure 17: The two components of stuttering. SMA = supplementary motor area.
The figure illustrates the condition of stuttering: you can’t continue, and you can’t stop. It may feel like a loss of control, but it is the conflict of two control systems. The two components of stuttering are: (1) the inhibition of a speech motor program because of an invalid error signal (left side) and (2) the speaker’s spontaneous attempt to continue talking (right side). The left side represents the behavior of the lateral premotor system, the right side that of the medial premotor system in Goldberg’s model. The left side is further equivalent to the causal chain depicted in Figure 7 (or see this version with explanations).
The two components depicted at the left and the right side of the figure are opposed in relation to the person’s will: The inhibitory response to an invalid error signal comes about against the person’s will, whereas the (over-) activation of the medial premotor system is the expression of the person’s will to continue talking. This two-component model is well consistent with the new insight provided by Neef et al. (2018) and Montag et al. (2019) because the person’s will is driven by the motivation to tell one’s thought, to express one’s emotion. Just this seems to be the reward of speaking. Most stutterers, including myself, know the feeling of disappointment when someone else in good faith completes the word or clause oneself doesn’t get out: We feel cheated out of our deserved reward. At first view it appears odd that a doing is its own reward, but just this seems to be the case not only in speaking, but also in singing, dancing, and in other joyful activities.
However, basal ganglia and medial premotor system seem to play a further role in stuttering: Metzger et al. (2018) found a correlation between substantia nigra activation and stuttering severity during a non-speech motor task (dopamine is released from the substantia nigra to the striatum). This as well as the results of non-speech behavioral studies (e.g., Markett et al., 2016) and the prevalence of hyperactivity among stutterers suggest that an overactive medial premotor system is a personality trait of not a few stutterers (see Section 2.8.1 in the main text.
The medial premotor system therefore seems to play a double role in stuttering. It is not only involved in the production of overt symptoms and influences their severity. The medial premotor system controls voluntary actions, and voluntary action is typically associated with goal-directed attention (note: goals are related to motivation and reward). A tendentially overactive medial premotor system may therefore supports an attentional imbalance associated with poor processing and reduced integration of sensory information in motor control (symbolized by the curved arrow in the figure). This seems to be a factor in the predisposition for developmental stuttering.
A core assumption of my theory is that stuttering is caused by insufficient processing of the auditory feedback of speech. The well-known fact that stuttering usually disappears with auditory masking as well as in mouthing (silent speech movements) can easily be explained: In these conditions, auditory feedback runs via the internal feedback loop, which seems to be automatically associated with sufficient processing such that stuttering does not occur (see Section 3.1 in the main text).
However, the processing of auditory feedback, it may run via the internal or the external loop, depends on proper functioning of the speech comprehension region in the left posterior superior temporal cortex, traditionally referred to as Wernicke’s area. The function of this cortical region is impaired in patients suffering of a Wernicke’s aphasia or receptive aphasia. They have difficulty understanding the speech of another person, but also in monitoring their own speech. The processing of auditory feedback is because of a structural damage of Wernicke’s area (e.g. after stroke). But if so, and if insufficient processing of auditory feedback causes stuttering – why do these patients not stutter,?
They indeed have difficulty in speech production: the words they are using do often not belong together or make sense (see, e.g. this video). But they do not have difficulty with producing connected speech that flows; hence Wernicke’s aphasia is also referred to as fluent aphasia. A theory claiming that stuttering is caused by insufficient processing of auditory feedback should be able to explain why a damage in Wernicke’s area is not associated with stuttering.
.
The fact that Wernicke’s aphasics are fluent seems to contradict my position that auditory feedback (via the external or the internal loop) and its proper processing is essential for speech fluency. But note that I don’t assume a feedback-based control of fluent speech; I rather assume that speech is mainly feed-forward controlled by motor programs of familiar syllables, words, and phrases (Section 1.1 and 1.2 in the main text). Those motor programs may be localized mainly in the left pre-motor area, such that their functioning is not impaired in Wernicke’s aphasia.
Auditory feedback is relevant for fluency only insofar as it forms the basis of the self-monitoring of speech, with error signals resulting in interruptions of speech flow. After my theory, poor processing of auditory feedback results in invalid error signals and thus in interruptions of speech flow without any error made by the speaker (Section 2.1).Why do such invalid error signals not occur in Wernicke’s aphasia, when auditory feedback is poorly processed in these patients’ brains?
The self-monitoring of speech encompasses a phonological, a syntactic, and a semantic component. The invalid error signals assumed to cause stuttering occur in phonological self-monitoring, the basis of which is a prediction of the correct sound sequence of a word or phrase. In Section 1.4 in the main text, I explain why these predictions or expectations cannot be generated on the basis of a motor plan (as the efference copy theory has proposed), but only on the basis of auditory feedback: The word or phrase is recognized on the basis of the initial phonemes (analysis-by-synthesis model), and this allows to predict its correct sequence. The internal monitor then compares this prediction with the further incoming feedback of this word or phrase, and any significant difference between prediction and perception elicits an error signal.
That means: Error signals can only occur, if the first step of the monitoring process – the recognition of the word and phrase produced – can take place. And already this seems not to be the case in Wernicke’s aphasia: they do not recognize the words they produce, thus no sound sequence of these words can be predicted. So there is no basis for self-monitoring and for error signals, neither valid nor invalid ones, and thus no basis for stuttering to occur.
Developmental stuttering, according to my theory, is a disorder in the automatic self-monitoring of speech. Stuttering can therefore not occur where this self-monitoring lacks. This seems to be the case in patients with a Wernicke’s aphasia: They don’t correct the many speech errors they make. They are not aware of them. And just this lack of self-monitoring is the cause why they are always fluent...