 A Theory of Stuttering
A Theory of Stuttering
A Theory of Stuttering
In normally fluent speakers, the cortical activity associated with speech processing is usually lateralized to the left hemisphere in almost all right-handers, but also in many left-handers. Therefore, the left brain hemisphere is referred to as language-dominant. Stuttering is typically associated with a lack of lateralization to the left hemisphere; activation is more symmetrical. This was suspected to be a cause of the disorder by some researchers in the 1920s (see, e.g., Travis, 1978). This is hardly consistent with the findings of Sowman et al. (2014). In an MEG study with stuttering and non-stuttering preschool children, they found no between-group differences in the lateralization of brain activity during picture-naming. In this section, I will propose that the lack of lateralization may result from a misallocation of attention during speech.
The idea that the lateralization of speech processing may be related to attention is not new. Kinsbourne (1970) proposed an attention model of lateralization, assuming that “the simple act of anticipation of verbal stimuli may preferentially activate the left hemisphere”. The attention model has been supported by several studies not only of brain activation (see below), but also of cochlear activity as a function of attention. Markevych et al. (2011) found a shared variance between central (cortical) and peripheral (cochlear) lateralization in the processing of consonant-vowel syllables, influenced by attention.
In fMRI studies showed that attention to the auditory channel strongly influenced the activation in auditory cortical areas. Hugdahl et al. (2003) reported that attention to vowels or words was associated with a leftward asymmetry in brain activation; passive listening was not. Sabri et al. (2008) found that some speech-related areas, particularly on the left hemisphere, were more active when verbal stimuli were presented, but only if participants listened to them, and not if they ignored them. Rämä et al. (2012) found greater activations in the right superior temporal gyrus when right-handed participants were distracted from auditory verbal stimuli, compared to a condition in which the participants listened to the stimuli.
Poeppel et al. (1996) found M100 responses (a component of the event-related brain potential, about 100 ms after stimulus onset) to be symmetric over both hemispheres when syllables were passively heard, but the response increased over the left temporal cortex and decreased over the right when participants actively listened to the syllables. According to the authors, the lateralized cortical response dependent on attention to verbal stimuli was concordant with hemispheric language dominance.
The lateralization of speech processing can be influenced even by whether attention is focused on the lexical or on the prosodic aspect of speech. Vingerhoets, Berckmoes, and Stroobant (2003) found activation more left-lateralized when participants paid attention to the lexical features of a sentence, but more symmetric with attention to the prosody (resulting from an increase on the right hemisphere).
Together, these findings suggest that active listening to verbal stimuli is associated with greater activation in the language-dominant hemisphere; passive hearing and ignoring verbal stimuli are associated with a more symmetric activation bilaterally. However, the studies reported above were conducted with normally fluent participants, not with stutterers, and they used speech recordings as stimuli, not the auditory feedback of speech.
A study with stutterers was conducted by Sato et al. (2011): children and adults who stutter and normally fluent controls listened to aurally presented verbal stimuli that contained either a phonemic or a prosodic contrast. In the controls, there was a clear left-hemispheric advantage for the phonemic contrast compared to the prosodic. Children and adults who stutter, by contrast, showed no significant difference between the two stimulus conditions. No one of the stutterers showed a left advantage when a phonemic contrast was presented.
However, Sato and colleagues did not ask their participants to pay attention to the stimuli or to specific features of them, as was the case in the studies with normal speakers mentioned above. A central assumption in my theory is that stutterers differ from non-stutterers in their habitual allocation of attention, such that they tend more towards passive listening. This would account for the lack of left advantage in the phonemic contrast condition because phonological processing hardly takes place without attention to the verbal stimulus (Cherry, 1953).
The study last mentioned was indeed conducted with stutterers, but the stimulus was not the auditory feedback of their speech. However, from neuroimaging studies, we know that cortical auditory association areas are active during speech in non-stutterers but not, or significantly weaker, in stutterers (see Table 1). I therefore assume that non-stutterers automatically allocate their attention so that their speech is perceived and processed properly, and mainly in the language-dominant hemisphere. In stutterers, by contrast, auditory feedback seems to be processed poorly, more like noise, which manifests in both, a lack of activation and, as a consequence, lateralization, in the temporal cortex.
A lack of left-lateralization in stutterers has not only been observed in the temporal cortex, but also during speech in the frontal cortex, mainly because of overactivity in the right frontal cortex (e.g., Budde, Baron, & Fox, 2014). For instance, the right frontal operculum (RFO) is the right-hemispheric homolog of Broca’s area. In normally fluent speakers, RFO is increasingly involved in speech processing when difficulties arise and usual routines no longer suffice (Mitchell & Crow, 2005; Taylor & Regard, 2003). It is not surprising that this brain region was found to be overactive in stutterers during speech (e.g., Neumann et al., 2003).
In sum, I think that the lack of lateralization to the language-dominant hemisphere in stutterers results from the misallocation of attention during speech. The lack of activation in the temporal cortex is caused by insufficient attention to auditory feedback, and the overactivation in the right frontal cortex results from compensatory behaviors, such as volitional speech-motor control and excessive speech planning.
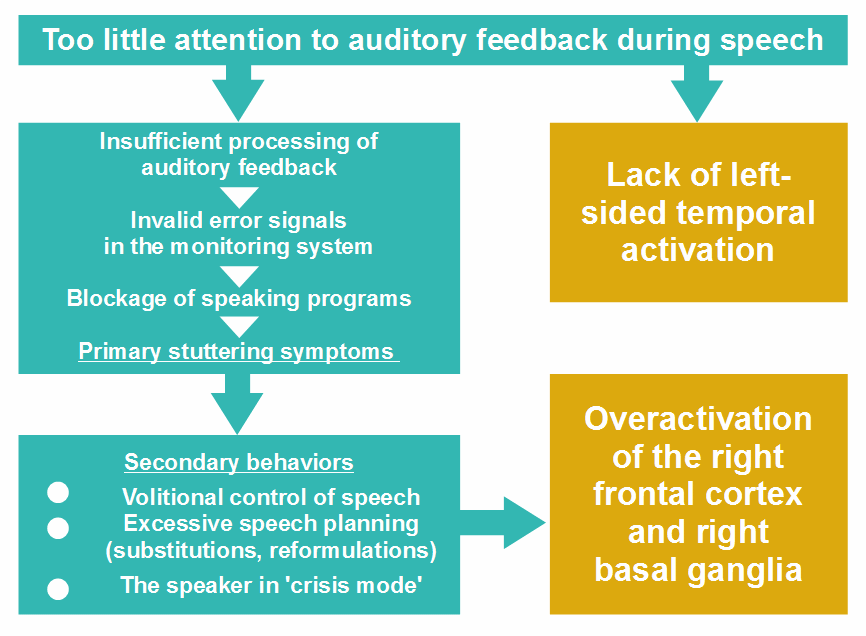
Figure 14: Misallocation of attention and the lack of lateralization of speech processing in stutterers. Note that some secondary behaviors take much attention, thereby reinforcing misallocation of attention (see Fig. ,10), but I have omitted the revertive arrow for the sake of clarity.
Some researchers hypothesized that the right-shift of brain activation observed in stutterers may compensate for structural or functional deficits in the left hemisphere; but the compensation is not satisfactory, and stuttering remains (e.g., Neumann et al., 2003). However, the idea that the brain compensates for a deficit in its functioning is problematic. There is no boss or supervisor in the brain who could notice that a part is not working properly and engage another part to take over that function. Instead, the person becomes aware of a problem and tries to cope with it by a change in behavior. A permanent change in behavior will result in a change in brain function and, with time, in brain structure (read more).
In this context, the results by Chang et al. (2011) are interesting. They investigated the structural and functional connectivity between several speech-related brain areas in adult stutterers and normally fluent controls. Results showed that between BA44 (part of Broca’s area) and pre-motor/motor areas, structural connectivity (i.e., fiber density) was lower on the left hemisphere in stutterers compared to controls, but this deficit was compensated for on the right hemisphere, where fiber density was higher in the stuttering group. By contrast, between BA44 and the posterior superior temporal gyrus (pSTG), structural connectivity on the left hemisphere was also much lower in the stutterer group compared to controls (see Fig. 4 in Chang et al., 2011), but this deficit, which may affect auditory-motor integration, was not compensated for on the right hemisphere.
In several studies, the structural integrity (fiber density) of white matter tracts in stutterers was compared with that in normally fluent speakers. In stutterers, as a group, deficits were also found in the corpus callosum (Chang et al., 2015; Civier et al., 2015; Connally et al., 2014; Cykowski et al., 2010). Corpus callosum fibers connect homologous areas of both cortical hemispheres and regulate the division of labor between them, with an area of one hemisphere inhibiting the activity of the homologous area of the other hemisphere. Hence, callosal fibers are crucial for the lateralization of speech processing, and the question arises whether callosal fiber deficits are causal for the lack of language lateralization or even for stuttering.
Szaflarski et al. (2006) examined 170 healthy right-handed children and adults aged between 5 and 67 years. They found that lateralization to the language-dominant hemisphere tended to increase between the ages of 5 and 20 years. This suggests that language lateralization is not innate (as language itself is not innate) but develops with language acquisition and the automation of speech-language abilities.
Furthermore, Chang et al. (2015) compared the structural integrity of white matter in young children aged between 3 and 10 years, with and without stuttering. They indeed found structural deficits in several parts of the corpus callosum in the stuttering group, but the group differences increased with age, that is, with the time the children had already stuttered, and the difference between the youngest children was minimal (see Chang et al., 2015, Fig. 2, lower row).
Putting the results of the two last-mentioned studies together, they do not support the hypothesis that the structural deficits in the corpus callosum in stutterers are causal for the lack of language lateralization or for stuttering itself. I rather assume that both, white matter deficits in the corpus callosum and the lack of language lateralization, are a result or concomitant of persistent stuttering. I agree with Civier et al. (2015), who point to a disadvantageous recruitment of the right frontal cortex in speech production, but I don’t believe that a reduced interhemispheric inhibition (due to the fiber deficits) is the cause.
I rather think that fibers are weaker because they were less frequently activated over time. Fiber maturation depends on the frequency of activation (Fields, 2010; Wake, Lee, & Fields, 2011; Zatorr, Fields, & Johansen-Berg, 2012; see also the next section). The maturation of the callosal fibers involved in language lateralization may thus depend on (progress in interaction with) the automation of speech production. Stuttering slows down the automation of speech production, and this may also slow down the fiber maturation in some parts of the corpus callosum.
Stuttering children, after having developed an awareness of the disorder, lose their confidence in the automatic control of speech—the confidence that their muscles do what they should do when speaking. Hence, they frequently are in a mode of crisis management when speaking, with the consequence that automatic speech control, including the inhibition of non-dominant speech areas, does not become well established and stabilized.
Demirel et al. (2024) analysed fMRI data from children and adults who stutter, and typically fluent speakers (119 participants in total) during four different speech and language tasks: overt sentence reading, overt picture description, covert sentence reading and covert auditory naming. They found no difference in laterality between stutterers and typically fluent speakers during the tasks. The result is surprising, but does not contradict my basic position that stuttering is not caused by altered language lateralization.
Changes in behavior entail changes in brain activation, and if they last over time, changes in brain activation result in structural changes: an increase or decrease in gray or white matter volume or fiber density. These effects were demonstrated in some studies (Bengtsson et al., 2005; Draganski et al, 2004; Driemeyer et al., 2008; Keller & Just, 2009; Scholz et al., 2009). That’s important for the therapy of stuttering, since almost all therapies aim to change the client’s behavior in some way.
(return)