 A Theory of Stuttering
A Theory of Stuttering
A Theory of Stuttering
If my assumptions about transient stuttering (in the preceding section) are correct, the question arises why a minority of the affected children don’t learn to adapt their attention to the requirements of connected speech. Two causal factors may be responsible: (1) an imbalance in the attention system, similarly as in transient stuttering but persistent, and (2) a deficit in auditory processing, which may not be present in children who spontaneously recover. Perhaps, both factors interact with one another. The development of persistent stuttering is sketched in the figure:
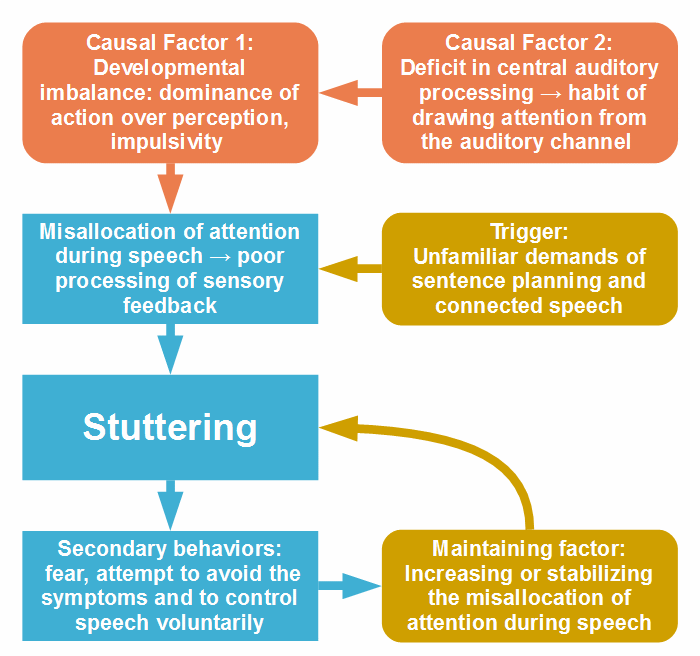
Figure 9: The development of persistent stuttering.
The idea that transient and persistent developmental stuttering are eddentailly the same disorder, and that persistence is caused by one or more additional factors has already been proposed by Ambrose, Cox, and Yairi (1997) in a study of the genetic basis of persistence and recovery. In the Abstract, they write: “It was found that recovery or persistence is indeed transmitted, and further, that recovery does not appear to be a genetically milder form of stuttering, nor do the two types of stuttering appear to be genetically independent disorders. Data are most consistent with the hypothesis that persistent and recovered stuttering possess a common genetic etiology, and that persistence is, in part, due to additional genetic factors.”
A finding by Chow and Chang (2017), distinguished the group of stuttering children who eventually persisted from the group that eventually recovered: an initially (in the youngest children) higher fractional anisotropy in the thalamic radiation and its abnormal, consistent or descending developmental trajectory (see Fig. 2, Clusters 8, 9, and 10 in the study). These findings can hardly be a consequence of stuttering, as the differences were greatest in the youngest children. Since the thalamus plays a central role in attention regulation, the findings may reflect an abnormal development of the attention system.
Furthermore, an anomalous functional connectivity between the attention networks and the default mode network (DMN) in children who eventually persisted in stuttering (Fig. 3 in Chang et al., 2018) indicates problems in the attention system in this group. These problems seem to partly differ from those in children with transient stuttering. Normally, the development proceeds so that the other brain networks segregate from the DMN with age. Therefore, the hyper-connectivity between the ventral attention network and the DMN in children with transient stuttering (mentioned in the last section) can be considered a transient delay in development. In children who persist in stuttering, by contrast, the connectivity between both, the ventral and the dorsal attention network, and the DMN is mostly reduced, suggesting a premature segregation of the networks in these children. This may have negative consequences for the further development of the attention system and its functioning in the control of behavior.
Moreover, in not a few behavioral studies, both, children and adults who stutter, exhibited deficits in attention regulation (see Section 3.3). Findings indicate that stutterers, on average, are less able to divide attention under dual-task conditions, less able to quickly shift attention and to suppress a planned motor action. For instance, Eggers, De Nil, and Van den Bergh (2012) concluded from their data that the orienting network, which is important for the allocation of attention, appears to be less efficient in children who stutter. Likewise, Kaganovich, Hampton Wray, and Weber-Fox (2010), who examined auditory processing in preschool-aged children who stutter, came to the conclusion that stuttering may be associated with less efficient attention allocation.
Further, stutterers are more prone to exhibit attention disorders (symptoms of ADHD, particularly hyperactivity and impulsivity; see Section 3.3). Interestingly, both, persistent developmental stuttering and ADHD, are more often diagnosed in males than in females (for ADHD, see Ramtekkar et al., 2010), which can be taken as a suggestion that a tendency towards hyperactivity, namely in the sense of a preponderance of action over perception, is responsible for the greater prevalence of persistent stuttering in males.
There are many findings of deficits in the processing of non-linguistic and linguistic acoustic stimuli in both, adults and children who stutter; see a list of relevant papers. The nature of the deficit is not yet clear; however, there are indications to problems in the interhemispheric corroboration of the auditory cortices (Salmelin et al., 1998; Kikuchi et al., 2017) and, probably resulting, a deficit in auditory gating, that is, in the suppression of redundant auditory input [Kikuchi et al., 2011; Saltuklaroglu et al., 2017. Auditory gating is fundamental for the control of auditory attention, and a deficit may lead to compensation at a higher level of attention control: to avoid acoustic over-stimulation, affected children may develop a habit of ignoring auditory input, except during active listening.
Unfortunately, there are few data about differences between those who persisted in stuttering and those who recovered in terms of auditory processing. The only study I know of is that by Howell, Davis, and Williams (2006). They compared children of both groups in a backward-masking task, the performance of which is assumed to reflect the operation of central auditory processing. They found an approximately 10 dB higher mean backward-masking threshold in the persistent group; that is, the probe tone needed to be about 10 dB louder on average for these children to hear it prior to the masking noise. The group difference was significant, but there was a high variety in the persistent group. The authors conclude that an auditory deficit may be sufficient, but not necessary, for the disorder to persist.
In a preceding study, Howell et al. (2000) had found a higher backward-masking threshold on average in stuttering children as compared to normal fluent controls, with that threshold being positively correlated with stuttering frequency in the stuttering group; that is, the poorer auditory processing, the higher stuttering frequency. A relationship between deficits in auditory processing and stuttering frequency or severity was also found in other studies (e.g., Beal et al., 2010, 2011; Jansson-Verkasalo et al., 2014; Kikuchi et al., 2017; Liotti et al., 2010). If a causal relationship exists between deficient auditory processing and stuttering, then it is unlikely that stuttering causes the auditory processing deficit. It is much more likely that the auditory processing deficit is a causal factor in stuttering.
Howell and Davis, 2011) concluded from their results that stuttering severity in young children predicts eventual recovery or persistence. If so, and if there is a correlation between stuttering severity and auditory processing (as was found in the studies mentioned in the preceding paragraph), then auditory processing in young stuttering children should predict persistence or recovery. This hypothesis is consistent with the results of Howell, Davis, and Williams (2006) reported above and can be tested in a longitudinal study.
Some further findings suggest that a deficit in auditory processing is the factor that distinguishes transient and persistent stuttering. Usler and Weber-Fox (2015) and Mohan and Weber (2015) found differences between children with persistent stuttering and those who recovered in the processing of acoustically presented linguistic stimuli. Chow and Chang (2017) found lower FA in the splenium (the posterior part of the corpus callosum) in children who eventually persisted in stuttering, but not in those who eventually recovered (Fig. 1, Cluster 4 in the study). The affected fibers probably connect bilateral temporal regions (Kuvazeva, 2013), and lower FA may reflect less effective inter-hemispheric labor division in auditory processing (callosal fibers often have inhibitory functions, such that activation in one hemisphere inhibits the homologous area in the other hemisphere).
The difference in FA in the mentioned Cluster 4 in Chow and Chang (2017) is great already with the youngest children, and there is not much overlap between the persistent group, on the one hand, and the recovered and the control group, on the other hand. Therefore, the structural deficit in the splenium can hardly be a consequence of stuttering (we can assume this only if a structural abnormality grows with the time the child has stuttered). Interestingly, Chow, Liu, Bernstein Ratner, and Braun (in an unpublished study, presented at the 2014 ASHA Convention) found a correlation between reduced FA in the splenium and stuttering severity in adults who stutter.
In adolescents and adults whose stuttering has become persistent, symptoms do typically not occur immediately at sentence onset, i.e., on the first syllable of the first word. The cause might be that, over time, almost all stutterers learn to plan sentences incrementally, such that less planning effort is required at sentence onset. However, stuttering still mostly occurs more in the initial portion than at the end of phrases, clauses, and sentences (read more), because, with incremental planning as well, more capacity for planning is needed in the initial part of these units than at their end.
Different from early childhood stuttering, in persistent stuttering, sentence planning is no longer the main cause for the misallocation of attention during speech. Caruso et al. (1994) have shown that stutterers produce more disfluencies under cognitive stress, and cognitive stress comes about not only by high demands in sentence planning, but also by the attempt to avoid or cover stuttering.
Some other factors come in addition: Stuttering frequently occurs on the salient words of a sentence, on long words and on words with high information load ((read more)). This pattern as well may be caused by a misallocation of attention. If a speaker, focuses only on the intended message and the salient, emphasized words so that his or her attention runs ahead to these words, then too little attention (processing capacity) may remain for auditory feedback at the end of the preceding ‘unimportant’ word. The auditory feedback of his word, e.g., a short, unstressed function word, is then poorly processed, a mismatch in the monitoring system occurs, and the resulting error signal blocks the motor program of the subsequent ‘salient’ word.
The fact that stressed words, long words, or words with a high information load are more likely to be stuttered than short, unstressed, ‘inconspicuous’ words is thus not due to specific features of those words, but rather because the speaker’s attention tends to be prematurely focused on them.
A typical feature of persistent stuttering is that anticipating a block triggers a block. After a child has become aware of his or her stuttering, speaking is often angst-burdened, and attention is automatically focused on the feared word. The focus on the threat then interferes with the processing of non-threat-related information (Compton, 2003; Ferneyhough et al., 2013; Koster et al., 2004); in our context, with the processing of auditory feedback. Note that not auditory feedback is the threatened information (a stutter block is inaudible), but (1) the memory of previous trouble with that word and (2) the inner perception of an inhibition of articulation, phonation, or breathing. An impending block is first mentally anticipated or internally felt, but not heard.
In not a few cases, stutterers pay much attention to the avoidance or control of stuttering, or to the listener’s reactions to symptoms. To avoid stuttering, they choose their words carefully, substitute words with feared initial sounds, or focus on articulatory or breathing movements and their volitional control. All this increases the misallocation of attention. A vicious circle, as depicted in Figure 9, develops, in which secondary behaviors increase stuttering severity.
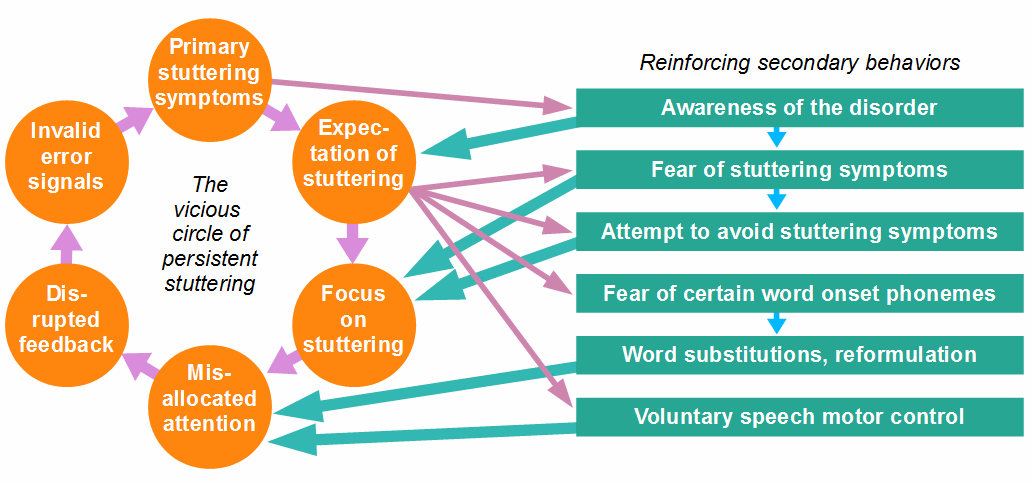
Figure 10: The vicious circle of persistent stuttering and the impact of some secondary behaviors.
Note that the causal chain (see Fig. 7) closes to a circle through the awareness of the disorder and the anticipation of stuttering events, and that attention allocation is the main interface through which secondary behaviors reinforce the vicious circle. However, as I have already written and justified above, I don’t assume that this vicious circle makes stuttering persistent; it only increases the frequency and severity of the symptoms.
In reading, the first word of a sentence is, on average, more frequently stuttered, even in older children and adults (Brown, 1945). However, Griggs and Still (1979) published individual results of a reading task, and as can be seen there, only three of their six subjects (four school-aged children and two adults) stuttered more frequently on the first word. In spontaneous speech, stuttering on the second, third, or fourth word of a sentence is typical for older children and adults, but stuttering on the first word is not (Heina, 1955, reported by St. Louis, 1979; Hannah & Gardner, 1968; Lanyon, 1969). Koopmans, Slis, and Rietveld (1991) found that 13.0% words on the first position and 15.6% words in the second position of clauses were stuttered on average in the spontaneous speech of adult stutterers.
I have often listened to stuttered speech in self-help groups and paid attention to whether symptoms occurred on the first words of sentences, and I found that it happened very rarely. This is not surprising because, on the one hand, the first word of an English or German sentence is typically a small function word (article, pronoun, preposition, etc.), and it is known that such words are less often stuttered than content words.
On the other hand, if a sentence starts with a content word, some stutterers have developed the habit of beginning with a ‘starter word’. This behavior indeed facilitates the start of the sentence, but because the speaker’s attention is already focused on the content word where he or she anticipates trouble, this content word is likely to be stuttered then.
(return)
Long words are more likely to be stuttered by adults (Brown & Moren, 1942; Quarrington, Conway, & Siegel, 1962; Schlesinger, Melkman. & Levi, 1966; Silverman & Williams, 1967; Soderberg, 1966; Taylor, 1966; Wingate, 1967) and less by children (Williams, Silverman, & Kools, 1969).
Information load refers to the predictability of a word within a context: the lower the predictability, the higher the information load. Words with a high information load are more frequently stuttered (Quarrington, 1965; Schlesinger et al., 1965; Soderberg, 1967). Rarely occurring words are more likely to be stuttered than frequently occurring words (Ronson, 1976; Schlesinger, Melkman, & Levi, 1966; Wingate, 1967); however, this effect overlaps with that of information load. See also St. Louis (1979) for an overview.
(return)